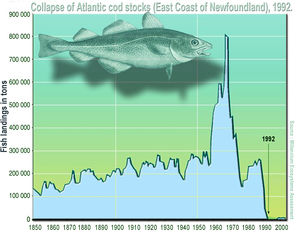
Tropical deforestation is driven primarily by frontier expansion of subsistence agriculture and large development programs involving resettlement, agriculture, and infrastructure. However, animal and plant population declines are typically pre-empted by hunting and logging activity well before the coup de grâce of deforestation is delivered. It is estimated that between 5 and 7 million hectares of tropical forests are logged annually, approximately 68-79% of the area that was completely deforested each year between 1990 and 2005 [FAO (Food and Agriculture Organization of the United Nations) 2007]. Tropical forests account for ~25% of the global industrial wood production worth US$400 billion or ~2% of the global gross domestic product [WCFSD (World Commission on Forests and Sustainable Development) 1998]. Much of this logging activity opens up new frontiers to wildlife and non-timber resource exploitation, and catalyses the transition into a landscape dominated by slash-and burn and large-scale agriculture. Few studies have examined the impacts of selective logging on commercially valuable timber species and comparisons among studies are limited because they often fail to employ comparable methods that are adequately reported. The best case studies come from the most valuable timber species that have already declined in much of their natural ranges.
For instance, the highly selective, but low intensity logging of broadleaf mahogany (Swietenia macrophylla), the most valuable widely traded Neotropical timber tree, is driven by the extraordinarily high prices in international markets, which makes it lucrative for loggers to open-up even remote wilderness areas at high transportation costs. Mechanized extraction of mahogany and other prime timber species impacts the forest by creating canopy gaps and imparting much collateral damage due to logging roads and skid trails (Grogan et al. 2008). Mahogany and other high-value tropical timber species worldwide share several traits that predispose them to commercial extirpation: excellent pliable wood of exceptional beauty; natural distributions in forests experiencing rapid conversion rates; low-density populations (often <1 tree/ha); and life histories generally characterized as non-pioneer late secondary, with fast growth rates, abiotic seed dispersal, and low-density seedlings requiring canopy disturbance for optimal seedling regeneration in the understory (Swaine and Whitmore 1988; Sodhi et al. 2008). One of the major obstacles to implementing a sustainable forestry sector in tropical countries is the lack of financial incentives for producers to limit offtakes to sustainable levels and invest in regeneration. Economic logic often dictates that trees should be felled whenever their rate of volume
increment drops below the prevailing interest rate (Pearce 1990). Postponing harvest beyond this point would incur an opportunity cost because profits from logging could be invested at a higher rate elsewhere.
This partly explains why many slow-growing timber species from tropical forests and savannahs are harvested unsustainably (e.g. East African Blackwood (Dalbergia melanoxylon) in the Miombo woodlands of Tanzania; Ball 2004). This is particularly the case where land tenure systems are unstable, and where there are no disincentives against ‘hit-and-run’ operations that mine the resource capital at one site and move on to undepleted areas elsewhere. This is clearly shown in a mahogany study in Bolivia where the smallest trees felled are ~40 cm in diameter, well below the legal minimum size (Gullison 1998). At this size, trees are increasing in volume at about 4% per year, whereas real mahogany price increases have averaged at only 1%, so that a 40-cm mahogany tree increases in value at about 5% annually, slowing down as the tree becomes larger. In contrast, real interest rates in Bolivia and other tropical countries are often >10%, creating a strong economic incentive to liquidate all trees of any value regardless of resource ownership. Tropical deforestation is driven primarily by frontier expansion of subsistence agriculture and large development programs involving resettlement, agriculture, and infrastructure. However, animal and plant population declines are typically pre-empted by hunting and logging activity well before the coup de grâce of deforestation is delivered.
It is estimated that between 5 and 7 million hectares of tropical forests are logged annually, approximately 68-79% of the area that was completely deforested each year between 1990 and 2005 [FAO (Food and Agriculture Organization of the United Nations) 2007]. Tropical forests account for ~25% of the global industrial wood production worth US$400 billion or ~2% of the global gross domestic product [WCFSD (World Commission on Forests and Sustainable Development) 1998]. Much of this logging activity opens up new frontiers to wildlife and non-timber resource exploitation, and catalyses the transition into a landscape dominated by slash-andburn and large-scale agriculture. Few studies have examined the impacts of selective logging on commercially valuable timber species and comparisons among studies are limited because they often fail to employ comparable methods that are adequately reported. The best case studies come from the most valuable timber species that have already declined in much of their natural ranges. For instance, the highly selective, but low intensity logging of broadleaf mahogany (Swietenia macrophylla), the most valuable widely traded Neotropical timber tree, is driven by the extraordinarily high prices in international markets, which makes it lucrative for loggers to open-up even remote wilderness areas at high transportation costs. Mechanized extraction of mahogany and other prime timber species impacts the forest by creating canopy gaps and imparting much collateral damage due to logging roads and skid trails (Grogan et al.
2008). Mahogany and other high-value tropical timber species worldwide share several traits that predispose them to commercial extirpation: excellent pliable wood of exceptional beauty; natural distributions in forests experiencing rapid conversion rates; low-density populations
(often <1 tree/ha); and life histories generally characterized as non-pioneer late secondary, with fast growth rates, abiotic seed dispersal, and low-density seedlings requiring canopy disturbance for optimal seedling regeneration in the understory (Swaine and Whitmore 1988; Sodhi et al. 2008). One of the major obstacles to implementing a sustainable forestry sector in tropical countries is the lack of financial incentives for producers to limit offtakes to sustainable levels and invest in regeneration.
Economic logic often dictates that trees should be felled whenever their rate of volume increment drops below the prevailing interest rate (Pearce 1990). Postponing harvest beyond this point would incur an opportunity cost because profits from logging could be invested at a higher rate elsewhere. This partly explains why many slow-growing timber species from tropical forests and savannahs are harvested unsustainably (e.g. East African Blackwood (Dalbergia melanoxylon) in the Miombo woodlands of Tanzania; Ball 2004). This is particularly the case where land tenure systems are unstable, and where there are no disincentives against ‘hit-and-run’ operations that mine the resource capital at one site and move on to undepleted areas elsewhere. This is clearly shown in a mahogany study in Bolivia where the smallest trees felled are ~40 cm in diameter, well below the legal minimum size (Gullison 1998). At this size, trees are increasing in volume at about 4% per year, whereas real mahogany price increases have averaged at only 1%, so that a 40-cm mahogany tree increases in value at about 5% annually, slowing down as the tree becomes larger. In contrast, real interest rates in Bolivia and other tropical countries are often >10%, creating a strong economic incentive to liquidate all trees of any value regardless of resource ownership.