Our rapacious appetite for both renewable and non - renewable resources has grown exponentially from our humble beginnings—when early humans exerted an ecological footprint no larger than that of other large omnivorous mammals— to currently one of the main driving forces in reorganizing the structure of many ecosystems. Humans have subsisted on wild plants and animals since the earliest primordial times, and most contemporary aboriginal societies remain primarily extractive in their daily quest for food, medicines, fiber and other biotic sources of raw materials to produce a wide range of utilitarian and ornamental artifacts. Modern hunter-gatherers and semi-subsistence farmers in tropical ecosystems, at varying stages of transition to an agricultural economy, still exploit a large number of plant and animal populations. By definition, exploited species extant today have been able to co-exist with some background level of exploitation. However, paleontological evidence suggests that prehistoric peoples have been driving prey populations to extinction long before the emergence of recorded history.
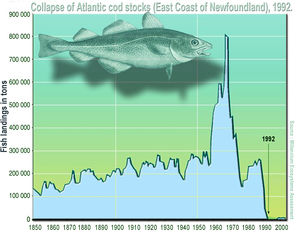
The late Paleolithic archaeology of big-game hunters in several parts of the world shows the sequential collapse of their majestic lifestyle. Flint spearheads manufactured by western European Cro - Magnons became gradually smaller as they shifted down to ever smaller kills, ranging in size from mammoths to rabbits (Martin 1984). Human colonization into previously people-free islands and continents has often coincided with a rapid wave of extinction events resulting from the sudden arrival of novel consumers. Mass extinction events of large-bodied vertebrates in Europe, parts of Asia, North and South America, Madagascar, and several archipelagos have all been attributed to post-Pleistocene human overkill (Martin and Wright 1967; Steadman 1995; McKinney 1997; Alroy 2001). These are relatively well corroborated in the (sub)fossil record but many more obscure target species extirpated by archaic hunters will remain undetected. In more recent times, exploitation-induced extinction events have also been common as European settlers wielding superior technology greatly expanded their territorial frontiers and introduced market and sport hunting. One example is the decimation of the vast North American buffalo (bison; Bison bison) herds. In the 1850s, tens of millions of these ungulates roamed the Great Plains in herds exceeding those ever known for any other mega herbivore, but by the century’s close, the bison was all but extinct.
Another example is the extirpation of mono dominant stands of Pau-Brasil legume trees (Caesalpinia echinata, Leguminosae- Mimosoidae) from eastern Brazil, a source of red dye and hardwood that gave Brazil its name. These were once extremely abundant and formed dense clusters along 3000 km of coastal Atlantic forest. This species sustained the first trade cycle between the new Portuguese colony and European markets and was relentlessly exploited from 1500 to 1875 when it finally became economically extinct (Dean 1996). Today, specimens of Pau-Brasil trees are largely confined to herbaria, arboreta and a few private collections. The aftershock of modern human arrival is still being felt in many previously inaccessible tropical forest frontiers, such as those in parts of Amazonia, where greater numbers of hunters wielding fire arms are emptying vast areas of its harvest sensitive mega fauna (Peres and Lake 2003). In many modern societies, the exploitative value of wildlife populations for either subsistence or commercial purposes has been gradually replaced by recreational values including both consumptive and non-consumptive uses. In 1990, over 20 million hunters in the United States spent over half a billion days afield in pursuit of wild game, and hunting licenses finance vast conservation areas in North America.
In 2006, ~ 87.5 million US residents spent ~US$122.3 billion in wildlife-related recreational activities, including ~US$76.6 billion spent on fishing and/or hunting by 33.9 million people (US Census Bureau 2006). Some 10% of this total is spent hunting white-tailed deer alone (Conover 1997). Consumptive uses of wildlife habitat are therefore instrumental in either financing or justifying much of the conservation acreage available in the 21st century from game reserves in Africa, Australia and North America to extractive reserves in Amazonia, to the reindeer rangelands of Scandinavia and the saiga steppes of Mongolia. Strong cultural or social factors regulating resource choice often affect which species are taken. For example, while people prefer to hunt large bodied mammals in tropical forests, feeding taboos and restrictions can switch “on or off” depending on levels of game depletion (Ross 1978) as predicted by foraging theory. This is consistent with the process of de-tabooing speciesthat were once tabooed, as the case of brocket deer among the Siona-Secoya (Hames and Vickers 1982). However, several studies suggest that cultural factors breakdown and play a lesser role when large-bodied game species become scarce, thereby forcing discriminate harvesters to become less selective (Jerozolimski and Peres 2003).